基于Ⅰ型内含子的体外环化策略是合成 circRNA 的主流方法。本期内容,我们将重点分享
Ⅰ型内含子的序列、结构及自我剪接机制
。
19 世纪 80 年代初期,TomasR. Cech 发现在体外没有蛋白质帮助的情况下,嗜热四膜虫
(Tetrahymena thermophila)
核糖体大亚基 26S rRNA 前体序列中间的一段干预序列(Intervening sequence,IVS)会被精准切除,发生 IVS 的环化和外显子序列的连接。这段干预序列后来被称为Ⅰ型内含子,是一类可催化自我剪切和外显子连接的 RNA 分子。
Ⅰ型内含子是含量最为丰富的拥有自剪接活性的内含子,数量已经超过 20000 个,广泛分布于自然界中。从细菌、真菌、原生生物及植物,到细胞核、线粒体及叶绿体,再到 mRNA、tRNA 及 rRNA 基因,你都可以发现它们的存在。
1
Ⅰ型内含子的序列信息
首先,我们来看看经常作为研究模型的 3 种Ⅰ型内含子序列。你会发现,Ⅰ型内含子在序列上相差很大,从几百个核苷酸到几千个核苷酸均有分布。
第一种是最早发现的
嗜热四膜虫Ⅰ型内含子
,
在其
26S rRNA 基因
中间存在一段 413bp 的内含子序列。

第二种,
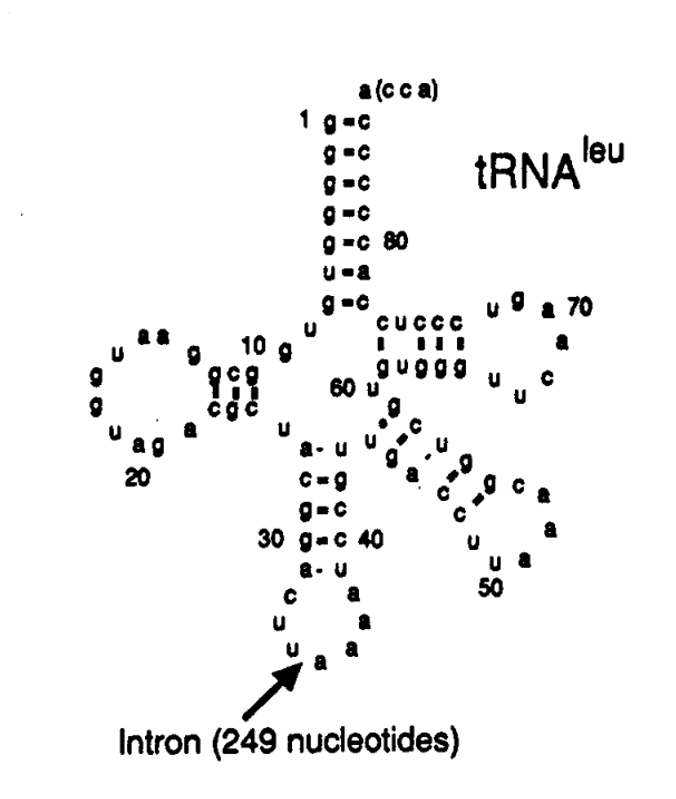
鱼腥藻 tRNA-Leu
基因中存在的一段 249bp 的Ⅰ型内含子序列。有意思的是,这段内含子序列位于 tRNA-Leu反密码子环中
(碱基 34 与 35 之间)
。

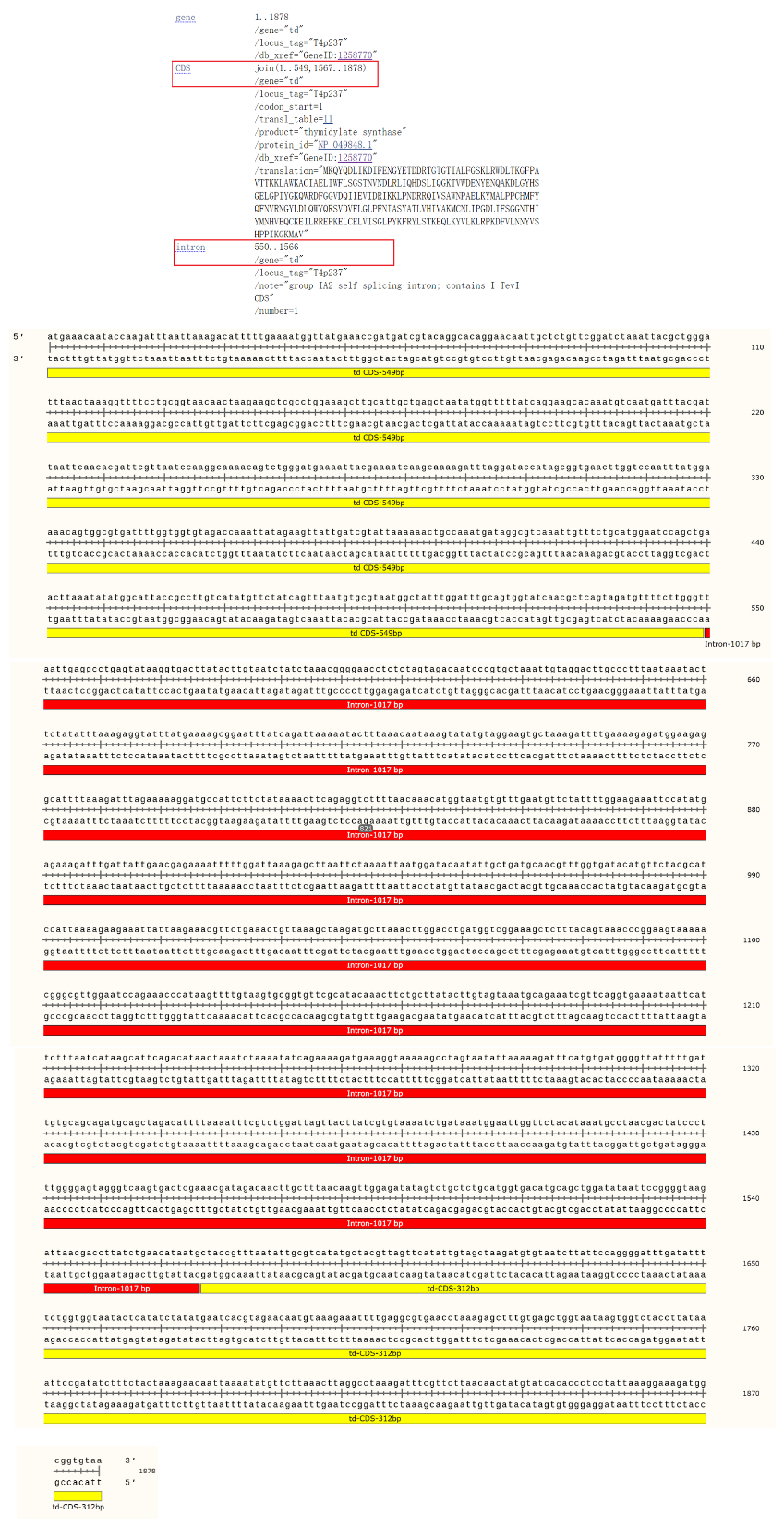
第三种,中断
T4 噬菌体胸苷合成酶基因
(thymidylate synthase,td)
的 1017bp 内含子序列,位于编码 GLY183 和编码 Leu184 三联体密码子之间。有意思的是,在 td 内含子序列中间检测到一个 735bp 的开放阅读框,以 5’端 AUG 起始,以 3’端 UAA 终止,实际上,这段 ORF 序列并能不翻译表达为蛋白。

2
Ⅰ型内含子的结构
2.1
要想理解Ⅰ型内含子的二级结构,首先得弄清楚这些二级结构的命名规则。1987 年,Henk F.Tabak 等人在 Nucleic Acids Research 发表文章:Structural conventions for group I introns,建立了Ⅰ型内含子二级结构的命名规则,旨在更好地促进自剪接型Ⅰ型内含子结构和功能的研究。
配对区,指的是Ⅰ型内含子二级结构中典型的碱基配对茎区
。根据 RNA 链 5'→3'方向,按照先后出现的顺序,依次命名为
P1~P9
。P2 是最靠近 P1 3'端的茎区,而 P9 是最靠近 P7 3'端的茎区。在某些内含子中,靠近 3'剪接位点的外显子序列与引导序列 IGS 之间形成互补配对区域,称为
P10
。有些内含子没有 P2,在这种情况下,配对区域依次命名为 P1,P3,P4 等。
位于保守配对区域 m 和 n 之间的额外茎区,根据 5'→3'方向,依次命名为 Pm1,Pm2 等,例如,在四膜虫Ⅰ型内含子中,在 P9 和 P10 之间存在的
P9.1,P9.2
。从保守配对茎区 m 延伸出去的茎区,根据 5'→3'方向,称为 Pma,Pmb,例如,
P5a,P5b,P6b,P9.1a
。如果配对区域的双链存在突出碱基,那么茎区 Pm 需要分隔为 a 和 b,例如,
P6a 和 P6b
。由于历史原因,P4 延伸出去的茎区称为 P5,而不是 P4a。位于配对区域 5'侧的配对碱基称为 Pm[5'],例如,在四膜虫Ⅰ型内含子中,
P4[5']
序列为 UGCGGG;位于配对区域 3'侧的配对碱基称为 Pm[3'],例如,
P4[3']
序列为 CCACGCA。
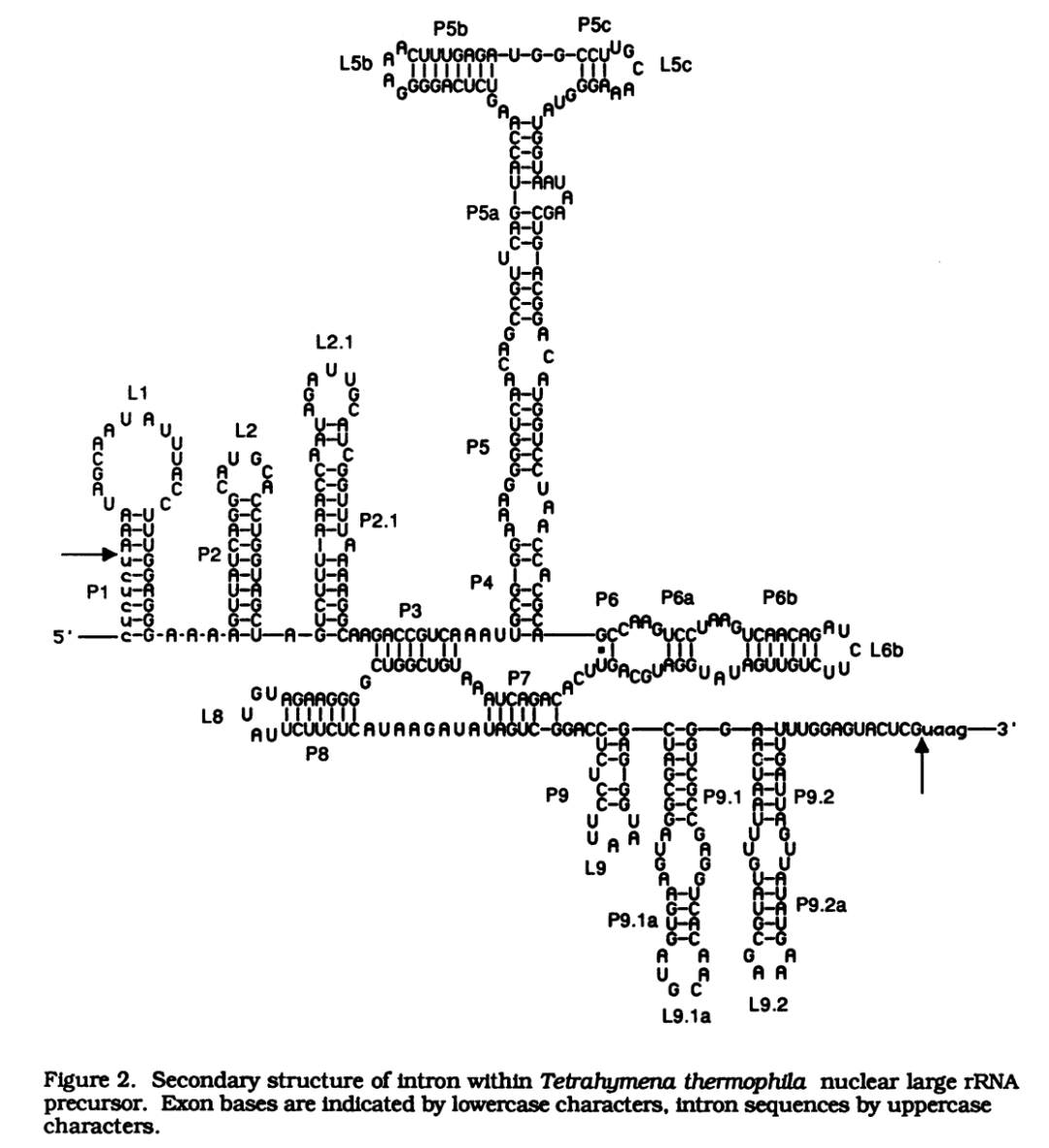
四膜虫大核糖体 Pre-rRNA 中的Ⅰ型内含子二级结构
Loop 环根据靠近他们的茎区命名,例如,
L1,L5b
等。需要注意的是,并非所有的配对区域都拥有 Loop 环。
连接区,指连接两个配对区的核苷酸。按照 5'→3'方向,根据他们连接的配对区命名。因此,在四膜虫Ⅰ型内含子中,把连接 P3 和 P4 的区域称为
J3/4
(AAU);把连接 P8 和 P7 的区域称为
J8/7
(AUAAGAUA)。
在Ⅰ型内含子中,存在 4 个保守的内部元件,分别称为
P,Q,R 和 S
。这些保守元件既有配对区,又有未配对区,例如,在四膜虫Ⅰ型内含子中,P 是由部分 J3/J4,所有的 P4[5']以及部分 J4/5 组成。
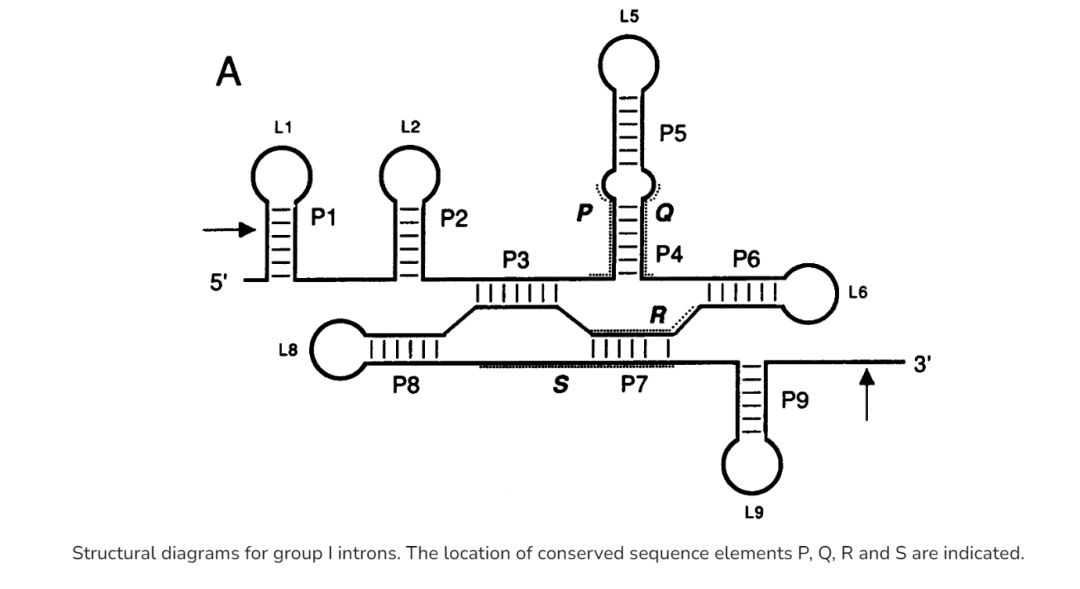
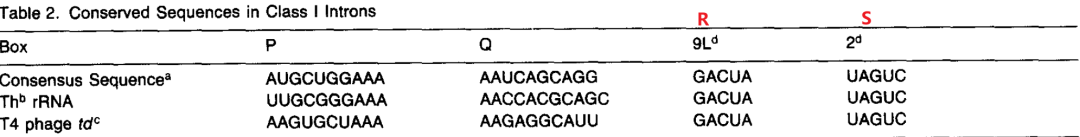
2.2
尽管 Ⅰ型内含子的一级序列并不高度保守,但是所有的Ⅰ型内含子都具有一个由 9 个保守的配对区
(P1~P9)
组成的高度保守的二级结构
。这 9 个配对区组成 3 个结构域:
P1~P2 结构域、P4~P6 结构域和 P3~P9 结构域
。P1 是底物配对区, 包含了核酶的 5′剪切位点。P3 和 P7 形成一个对核酶催化至关重要的假节结构。核酶的三级结构紧密折叠, 其中 P4~P6 结构域充当骨架, 而 P3~P9 结构域紧密包裹在 P4~P6 结构域的周围。核酶的 3 个结构域由保守的连接区连接, 这些连接区在核酶整体结 构的组装以及活性中心的形成中起到重要的作用,例如 J3/4 和 J6/7 可以同 P4~P6 结构域形成碱基三联配, 促进 P4~P6 与 P3~P9 之间的组装。
除了这 9 个保守的配对区外, 大多数 I 型内含 还具有至少一个额外的结构域, 被称为
周边结构
, 例如四膜虫内含子中的 P5abc, P2.1 和 P9.1-P9.2 等。这些周边结构主要是通过形成各种三级相互作用稳定核酶的核心结构, 它们的缺失总体来讲会降低但不会消除核酶的剪接活性,但是某些周边结构对核酶的活性却是必需的。
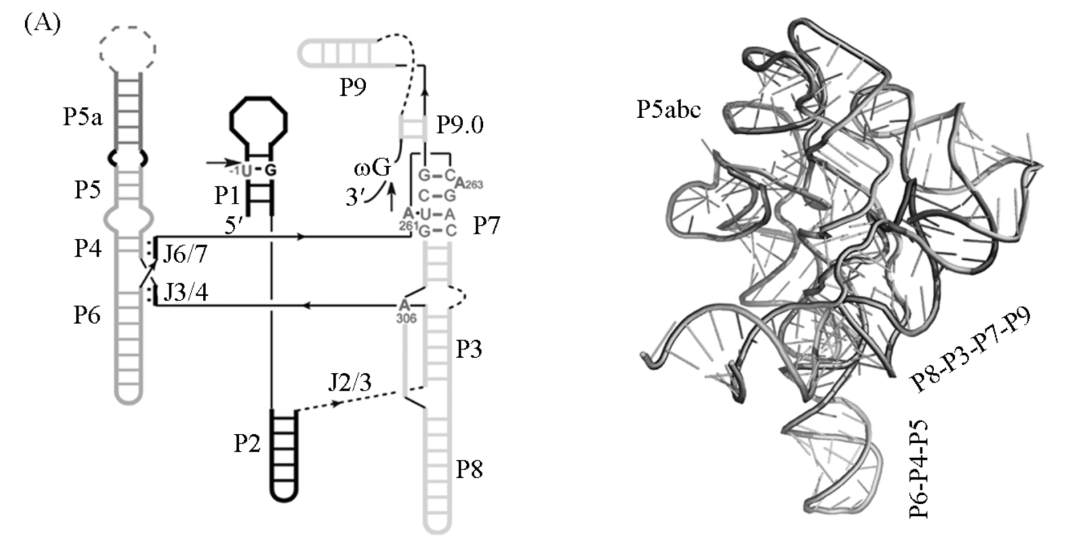
Ⅰ型内含子的二级结构和三级结构
2.3
与四膜虫大核糖体 Pre-rRNA 中的Ⅰ型内含子二级结构相比,T4 噬菌体胸苷酸合成酶基因中的Ⅰ型内含子结构中存在 P7.1 和 P7.2,但是,不存在 P2.1,而鱼腥藻 Pre-tRNA-leuⅠ型内含子中不存在 P7.1 和 P7.2,也不存在 P2.1 或者 P5abc,并且拥有一个非常特别的 P9。
在四膜虫大核糖体 Pre-rRNA 中的Ⅰ型内含子中组成 P1 的碱基对为
6 个
,在鱼腥藻 Pre-tRNA-leuⅠ型内含子中组成 P1 的碱基对
只有 3 个
,在 T4 噬菌体胸苷酸合成酶基因中的Ⅰ型内含子组成 P1 的碱基对为
7 个
。
在四膜虫大核糖体 Pre-rRNA 中的Ⅰ型内含子中组成 P10 的碱基对为
7 个
,在 T4 噬菌体胸苷酸合成酶基因中的Ⅰ型内含子中组成 P10 的碱基对为
2 个或者 3 个
,在鱼腥藻 Pre-tRNA-leuⅠ型内含子中组成 P10 的碱基对为
2 个或者 1 个。
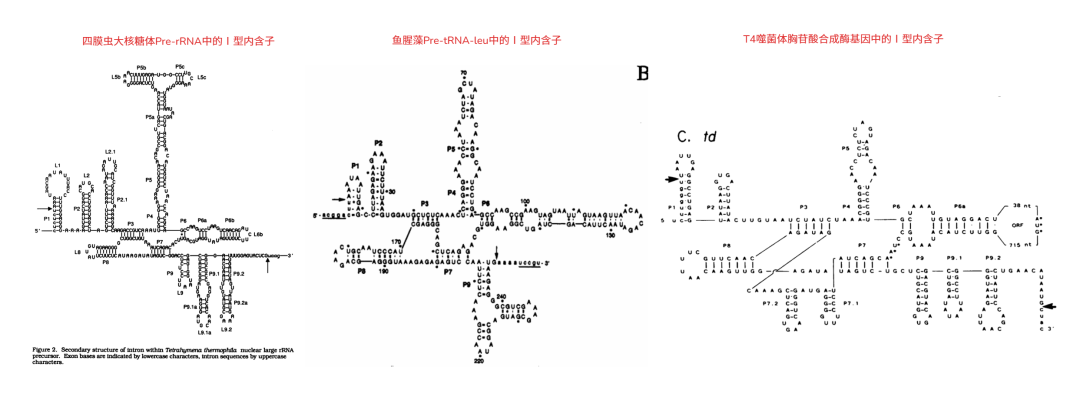
3
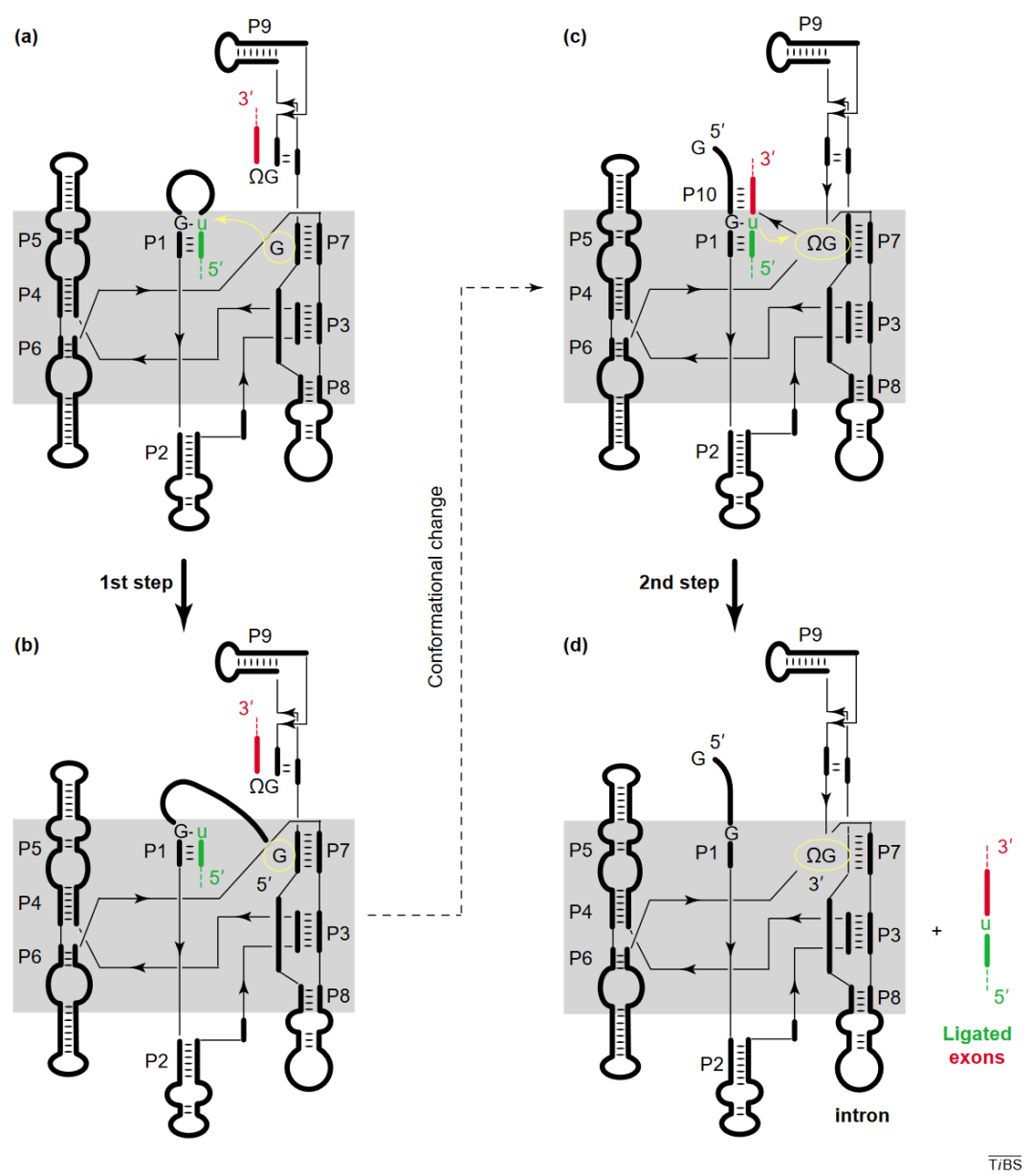
Ⅰ型内含子的自我剪接反应是一个连续的两步转酯反应。
5’外显子与引导序列 IGS 形成底物茎区 P1。核酶的 5’剪切位点由 P1 茎区中的保守配对
G•U
构成。在第一次转酯反应中,外源鸟苷酸被结合到内含子的活性中心,用其 3′羟基作为亲核试剂 ,攻击 5′剪切位点尿苷酸(U)的磷原子。该反应导致 5′剪切位点处的磷酸二酯键断裂,在 5′外显子末端露出游离的 3′羟基, 并使外源的鸟苷连接到内含子的 5′端。
内含子 3’末端是一个保守的鸟苷酸ΩG,决定 3’剪接位点。在第一次转酯反应完成后,内含子构象发生改变,使得
ΩG
进入活性中心接受来自 5′外显子末端游离 3′羟基的攻击,触发外显子的连接和内含子的释放。有两个非常重要的元件将 3’剪接位点的ΩG 拉到距离活性中心 P7 很近的位置:第一,由 P7 下游序列和内含子末端ΩG 上游的序列配对;第二,由 3′外显子和部分 IGS 序列形成的 P10。
识别微信二维码,添加生物制品圈小编,符合条件者即可加入
本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
到此这篇crrna是什么(crRNa是什么)的文章就介绍到这了,更多相关内容请继续浏览下面的相关推荐文章,希望大家都能在编程的领域有一番成就!
版权声明:
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。
如若内容造成侵权、违法违规、事实不符,请将相关资料发送至xkadmin@xkablog.com进行投诉反馈,一经查实,立即处理!
转载请注明出处,原文链接:https://www.xkablog.com/rfx/51156.html
